
1,
ライフサイエンス基盤研究
●
リボソーム30SサブユニットとRimMとの複合体およびリボソーム30SサブユニットとRNAとの複合体
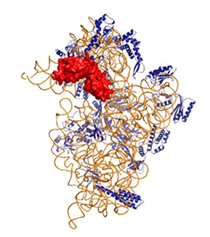
●
DEAD-box タンパク質 Vasa
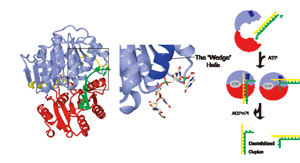
DEAD-boxタンパク質は幅広く保存されたRNAヘリケースのファミリーであり,ATPの加水分解を伴ってRNAの高次構造変化を引きおこす.転写,スプライシング,翻訳などのRNAが関与する様々な生命現象が,DEAD-boxタンパク質によって制御されている.
生殖細胞の分化を制御するDEAD-boxタンパク質であるVasaとRNA,ATPアナログからなる複合体の結晶構造を決定した.これは,DEAD-boxタンパク質が働く瞬間の構造を捉えた初めての例である.結晶構造中でVasaはRNAと密接に相互作用しており,保存されたα-ヘリックスを楔のように用いて結合RNAの鋭い折れ曲がりを誘起していた.そのような湾曲したRNAは塩基対を連続して形成できず,これによってRNA高次構造の変化が引き起こされていると考察される.すなわち,DEAD-box タンパク質は標的RNA部位に直接結合して湾曲を引き起こすことにより,複雑なRNA構造の特異的な制御を行っていると考えられる.
生殖細胞の分化を制御するDEAD-boxタンパク質であるVasaとRNA,ATPアナログからなる複合体の結晶構造を決定した.これは,DEAD-boxタンパク質が働く瞬間の構造を捉えた初めての例である.結晶構造中でVasaはRNAと密接に相互作用しており,保存されたα-ヘリックスを楔のように用いて結合RNAの鋭い折れ曲がりを誘起していた.そのような湾曲したRNAは塩基対を連続して形成できず,これによってRNA高次構造の変化が引き起こされていると考察される.すなわち,DEAD-box タンパク質は標的RNA部位に直接結合して湾曲を引き起こすことにより,複雑なRNA構造の特異的な制御を行っていると考えられる.
PDBj:
2DB3
PubMed:
16630817
●
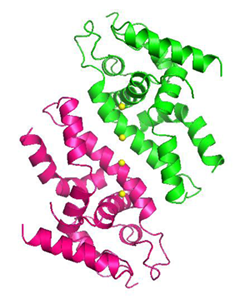
有鬚動物由来細胞外巨大へモグロビン

有鬚(ゆうしゅ)動物は口や消化管を持たず,かわりに体内に共生させた細菌がつくりだす有機物を栄養源にしていると考えられている.有鬚動物は分子量が数十万から数百万の,巨大な細胞外ヘモグロビン複合体を有しており,これにより宿主が利用する酸素と,共生細菌が利用する硫化水素を同時に結合・運搬するという,極めて特徴的な機能を発現している.
この巨大へモグロビン(分子量38万)の結晶構造を決定し,ヒトのヘモグロビンとはまったく異なる様式で4種類のサブユニットが会合し,全体として内部に大きな空洞のある,球状の24量体を形成していることを明らかにした.得られた結晶構造から硫化水素結合部位を推定し,その周辺環境から結合のメカニズムを議論した.これらの成果から,無脊椎動物のもつヘモグロビン複合体についての進化上の考察にも重要な知見となることが期待される.
この巨大へモグロビン(分子量38万)の結晶構造を決定し,ヒトのヘモグロビンとはまったく異なる様式で4種類のサブユニットが会合し,全体として内部に大きな空洞のある,球状の24量体を形成していることを明らかにした.得られた結晶構造から硫化水素結合部位を推定し,その周辺環境から結合のメカニズムを議論した.これらの成果から,無脊椎動物のもつヘモグロビン複合体についての進化上の考察にも重要な知見となることが期待される.
PubMed:
16204001
●
Dicer のRNase IIIb ドメイン
RNAi (RNA interference)は,dsRNA (double-stranded RNA)によって配列特異的な遺伝子抑制が引き起こされる現象である.RNAi反応過程では,Dicerが細胞内に導入されたdsRNAを21-24塩基からなるsiRNA (short interfering RNA)にプロセッシングする.siRNAは,RISC (RNA-induced silencing complex)に取り込まれ,siRNAと相補的なmRNAを認識し分解する.Dicerはマルチドメインからなるタンパク質で,N末端からヘリカーゼ,PAZドメイン,二つのタンデムなRNase IIIドメイン,dsRBD (dsRNA-binding domain)で構成される.
本研究では,ヒトDicerのC末端RNase IIIドメイン (RNase IIIb)の結晶構造を明らかにした.これまで構造が明らかにされていた,原核生物,下等真核生物由来のRNase IIIドメインとは異なる特徴を見出すことができた.Dicerは,転写制御を行う内在性のmiRNA (microRNA)の生産にも関与するため,この構造情報は発生・分化を理解する上で重要な意味を持つと考えられる.
本研究では,ヒトDicerのC末端RNase IIIドメイン (RNase IIIb)の結晶構造を明らかにした.これまで構造が明らかにされていた,原核生物,下等真核生物由来のRNase IIIドメインとは異なる特徴を見出すことができた.Dicerは,転写制御を行う内在性のmiRNA (microRNA)の生産にも関与するため,この構造情報は発生・分化を理解する上で重要な意味を持つと考えられる.
PDBj:
2EB1
PubMed:
17920623
●
ヒトCdc42-interacting protein 4(CIP4) extended FCH(EFC)ドメインとFormin-binding protein 17(FBP17)のEFC ドメインの結晶構造
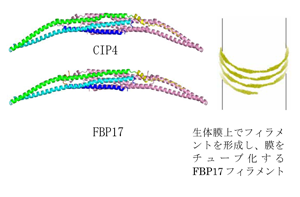
ヒトCIP4やFBP17等のEFCドメインを保持するタンパク質は,エンドサイトーシスや細胞分裂等の膜変形を伴う現象への関与が示唆されていた.これらのタンパク質のEFCドメインの結晶構造を決定したところ,EFCドメインは長く伸びた弓型の二量体を形成しており,弓型の内側が生体膜との結合や,生体膜のチューブ化に重要であることが分かった.タンパク質による生体膜のチューブ化には,タンパク質の生体膜上でのフィラメント化が重要な役割を果たす場合があることが示唆されてきた.実際,FBP17のEFCドメインはフィラメントを形成することが報告されている.今回の構造決定により,ヒトCIP4やFBP17のEFCドメインが,結晶中で類似のフィラメント構造を形成することを確認した.これに基づき,EFCドメインによる生体膜チューブ化の推測モデルを提案することが出来た.
PubMed:
17512409
●
糖タンパク質輸送受容体 (Emp46p,Emp47p)
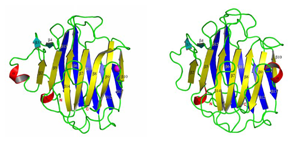
小胞体で合成された分泌経路を通る新生タンパク質は,N-結合型糖鎖の付加といった翻訳後修飾を受けた後,輸送小胞に乗せられて送り出される.これらのタンパク質(積み荷)の選別は,糖タンパク質輸送受容体によって行われていることが明らかになりつつある.本研究では,糖タンパク質輸送受容体(Emp46p,Emp47p)の糖鎖認識機構および糖タンパク質の輸送機構を明らかにすることを目的として,糖鎖認識ドメイン(CRD)単体およびそれらの金属結合体の結晶構造解析を行った.その結果,Emp46pおよびEmp47pは,同様の機能を担うカルシウム依存性ERGIC-53とは異なり,新しいカルシウム非依存性の糖タンパク質輸送受容体であることが明らかになった.糖タンパク質の輸送について新たな知見を与えた点で学術的価値は極めて高い.
PubMed:
16439369
●
記憶・学習に関与する糖転移酵素の構造解析
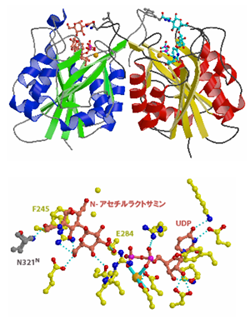
HNK-1糖鎖抗原は,神経系に存在する細胞接着因子や糖脂質に特徴的に発現し,神経系で重要な役割を果たしている.我々は,HNK-1糖鎖の生合成に必須なグルクロン酸転移酵素GlcAT-PおよびGlcAT-SのX線結晶構造解析を行った.その結果,他のグルクロン酸転移酵素では保存されていないアミノ酸が,受容体基質の分子識別を行っている機構を明らかにした.また,構造解析に基づいて作成した変異体は,GlcAT-PおよびGlcAT-Sの基質特異性を変換させることに成功した.どのような機構でHNK-1糖鎖の合成が行われているかを構造生物学的に明らかにした点で学術的価値が高いと共に,最終的には記憶や学習と糖鎖との関連を明らかにする研究の礎となるものである.
●
オートファジーを制御するタンパク質群の構造生物学
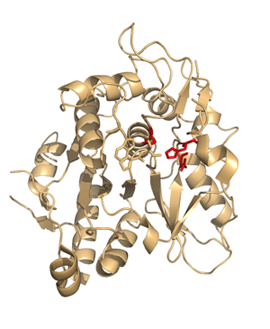
オートファジーとは,細胞質の一部をオートファゴソームと呼ばれるオルガネラで取り囲み,分解コンパートメントである液胞/リソソームへと輸送し,分解を行うシステムである.出芽酵母では,少なくとも16種類のAtg蛋白質群がオートファゴソームの形成に関わっていることがわかっているが,実際にどのような働きで形成されるのか,その分子機構についてはよくわかっていない.我々はAtg蛋白質群の三次元構造を網羅的にかつ様々な複合体として解析し,これら機能未知蛋白質群が互いにどのように連携を取り合って,オートファゴソームの形成に働いているのかを解明することを試みている.複数の種由来のAtgタンパク質を用いることで,現在までにAtg3,Atg4,Atg5,Atg8,Atg12,Atg16N末端領域の結晶構造の決定に成功した.現在それぞれの因子について構造に基づいた機能解析を進めている.オートファジーは抗原提示や神経変性疾患との関連も示されており,今後医学的にも重要な意義を持つものである.図はシステインプロテアーゼAtg4のヒトホモログ(HsAtg4B)の構造を示す.活性部位はループにより自己阻害されており,その活性が厳密に制御されている.
●
HLA-G ダイマー
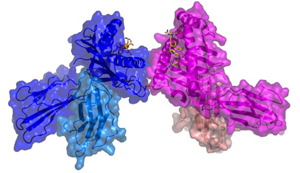
母体と胎児の接点である胎盤では免疫系の働きを抑える特殊な免疫寛容システムが存在する.胎盤の栄養膜細胞にはHLA-Gと呼ばれる細胞表面抗原が特異的に発現し,母体側の免疫細胞はHLA-Gに対する抑制性レセプター(LILRなど)を発現するため,胎盤では免疫寛容が誘導される.本研究では,HLA-Gが通常のモノマー型以外にダイマー型を有することに着目し,その機能解析を行った.HLA-Gはダイマー化により抑制性LILR群を介するシグナル伝達が100倍程度増強されることを見出した.さらに,HLA-Gダイマーの立体構造解析に成功し,HLA-Gがダイマー化によりLILRの細胞内ドメインが近接し,シグナルが増強される構造であることがわかり,機能解析を裏付ける結果であった.これらの結果はHLA-Gダイマーが免疫抑制タンパク製剤として応用可能であることを示すもので,特許申請を行った.
PDBj:
2D31
PubMed:
16455647
●
hnRNP D
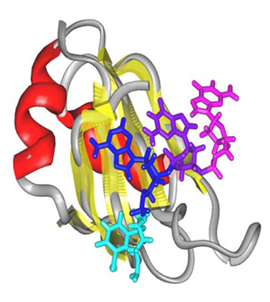
hnRNP Dタンパク質はテロメア配列のDNAに特異的に結合し,またテロメラーゼとも直接結合する.このためテロメアDNA長の制御への関与が示唆されており,老化・癌化とも関連している可能性がある.
本研究ではhnRNP Dタンパク質とテロメアDNAの複合体の構造を決定し,1本鎖状態のテロメアDNAがhnRNP Dによって特異的に認識されるメカニズムを解明した.またテロメアDNAは元々は4重鎖構造を形成しており,この構造がhnRNP Dによって1本鎖構造に遷移することも見出した.hnRNP D及び類縁のhnRNP A1タンパク質によるDNAの4重鎖から1本鎖への構造変換によって,DNAの複製の円滑な進行が保障されていることを,生化学的に示した.さらにこの構造変換が,テロメア長の伸長の保障にも寄与していることを示唆する細胞生物学的な結果も得つつある.hnRNP Dの働きを抑えることでテロメア長の短小化を引き起こし,癌に対する治療法につながることが期待される.
本研究ではhnRNP Dタンパク質とテロメアDNAの複合体の構造を決定し,1本鎖状態のテロメアDNAがhnRNP Dによって特異的に認識されるメカニズムを解明した.またテロメアDNAは元々は4重鎖構造を形成しており,この構造がhnRNP Dによって1本鎖構造に遷移することも見出した.hnRNP D及び類縁のhnRNP A1タンパク質によるDNAの4重鎖から1本鎖への構造変換によって,DNAの複製の円滑な進行が保障されていることを,生化学的に示した.さらにこの構造変換が,テロメア長の伸長の保障にも寄与していることを示唆する細胞生物学的な結果も得つつある.hnRNP Dの働きを抑えることでテロメア長の短小化を引き起こし,癌に対する治療法につながることが期待される.
PDBj:
1X0F
PubMed:
15734733
●
SUMO1-TDG 複合体
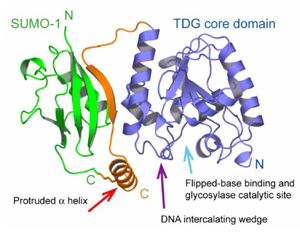
SUMO (Small ubiquitin like modifier)はユビキチン類似蛋白質の一つであり,ユビキチン化とよく似た一連の酵素反応により標的蛋白質のリジン残基を翻訳後修飾する.配列上は,SUMO-1はユビキチン(Ub)と18%の相同性しか示さないが,Ubに類似した立体構造を有している.SUMOは標的蛋白質の核輸送,転写制御,染色体分離など様々な核内現象に関与している.遺伝子修復酵素ヒトTDGは,単体では酵素のターンオーバー速度が極めて遅いため,効率よく遺伝子を修復し得ないが,SUMO化によりDNAから解離しやすくなって見かけの酵素活性があがる.そのメカニズムは長らく未知であったが,本研究により,SUMOと共有結合したTDGの結晶構造を解析することで,機構が解明された.すなわち,TDGには共有結合部位のほかにもう一箇所SUMOと分子間βシートを形成する部位が存在し,その二箇所に挟まれたTDGのループ部分に新しいヘリックス構造が誘起され,これがTDGに結合したDNAを押し出すアームとして機能する.このような翻訳後修飾にともなう構造の誘起の機構の発見は世界最初である.
PDBj:
1WYW
PubMed:
15959518
●
タンパク質合成調節因子/阻害タンパク質複合体
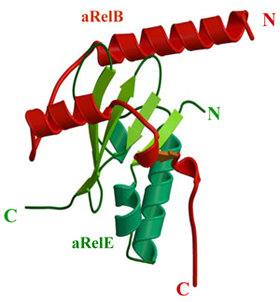
大腸菌に代表される真正細菌は,アミノ酸欠乏条件下でタンパク質合成反応を抑制しアミノ酸プールの減少を抑える調節因子・トキシン/アンチトキシン(RelE/RelB)を有している.即ち,遺伝子relBEはオペロンを形成し,relEはトキシン(RelE)を,relBはRelEの阻害タンパク質・アンチトキシン(RelB)をコードしている.通常,RelBはRelEと複合体(RelB-RelE)を形成することによりRelEの毒性を抑制している.しかし,アミノ酸欠乏下では細胞内プロテアーゼ・Lon等の細胞内プロテアーゼが活性化され,RelBのみが特異的に分解されることによりRelE が活性化される.その結果,RelEはリボソームのA部位に結合し,翻訳過程のmRNAを特異的に切断することによりタンパク質合成反応を抑制する.我々は,超好熱古細菌(Pyorococcus horikoshii)ゲノム中にrelB/relE相当遺伝子(PHS014/PHS013)を見出し,それらの大腸菌内での共発現後,組み換えタンパク質複合体(aRelB-aRelE)を分離精製した.さらに,精製したaRelB-aRelE複合体の結晶化に成功し,その結晶構造を2.3Åの分解能で決定した.その結果,aRelBは3本のαへリックスからなる伸びた構造を持つタンパク質で,aRelEは3本のαへリックスと5本のβストランドからなる球状タンパク質で,両タンパク質とも新規な構造を有していた.また,その結合様式はaRelBがaRelEの回りを取り囲むように複合体を形成していることを明らかにした.さらに,aRelEと伸長因子EF-Gとの構造比較から,aRelEがtRNAのアンチコドンループの分子擬態としてリボソームのA部位に結合することを強く示唆した.
PDBj:
1WMI
PubMed:
15768033
●
RNA ポリメラーゼ/ppGpp 複合体
RNAポリメラーゼは,DNAの遺伝情報を正確に読み取り,RNAを合成する重要なタンパク質である.緊縮調節と呼ばれる現象が知られており,細胞がアミノ酸の飢餓状態にさらされると,グアノシン4リン酸(ppGpp)と呼ぶ化学物質をつくり,RNAポリメラーゼの活性を制御し,特定の遺伝子の発現を抑制したり促進したりすることが分かっている.
本研究では,ppGppとRNAポリメラーゼの複合体を結晶状態で得ることに成功し,その結晶構造を決定した.その結果,ppGppは,RNAポリメラーゼの活性中心の立体配置に影響を及ぼし,DNAとも直接相互作用することによって,ポリメラーゼの活性を制御していると考えられる.転写反応とその制御に関わるメカニズムが原子レベルで解明されたことは,遺伝情報の伝達の仕組みを解する上で重要な知見を与えるだけでなく,効果の高い抗生物質の開発も可能にすると期待できる.
本研究では,ppGppとRNAポリメラーゼの複合体を結晶状態で得ることに成功し,その結晶構造を決定した.その結果,ppGppは,RNAポリメラーゼの活性中心の立体配置に影響を及ぼし,DNAとも直接相互作用することによって,ポリメラーゼの活性を制御していると考えられる.転写反応とその制御に関わるメカニズムが原子レベルで解明されたことは,遺伝情報の伝達の仕組みを解する上で重要な知見を与えるだけでなく,効果の高い抗生物質の開発も可能にすると期待できる.
PDBj:
1SMY
PubMed:
15109491
●
EF-P修飾酵素 GenX
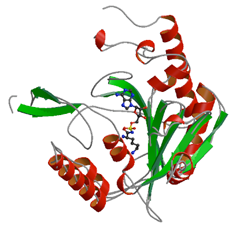
GenXは真正細菌のみに存在し,タンパク3000において構造解析を行った当時は機能が不明であった.配列はアミノアシルtRNA合成酵素(aaRS)のひとつであるLysRSに似ているが,アミノ酸であるリジンをtRNAに結合させるアミノアシル化活性は無いことが知られていた.その後,ターゲットタンパク研究プログラムにおけるGenXとEF-Pについての生化学的な実験により,EF-PがGenXの基質であることが発見された.また,GenX・EF-P複合体の立体構造にも成功し,その構造がaaRS・tRNA複合体の構造と酷似していることが判明した.さらに,aaRSがtRNAにアミノ酸を結合させる際と酷似した方法で,GenXがEF-Pへとアミノ酸を結合させていることを解明した.
PDBj:
3A5Y
PubMed:
20729861
●
リボソーム30Sサブユニットと成熟因子RbfAタンパク質との複合体
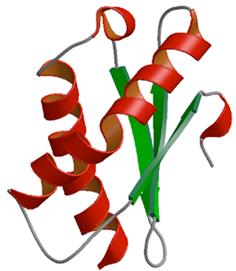
リボソーム30Sサブユニット(分子量約90万)は,メッセンジャーRNA(mRNA)に転写された遺伝情報を正確に読み取る役割があり,約1,500塩基のリボソームRNA(16S rRNA)と22個のリボソームタンパク質で構成されている.このサブユニットの成熟過程で重要な役割を果たしているタンパク質の一つがRbfAだが,RbfAの結合部位や結合の様式,またその役割の詳細については不明であった.
本研究では,高度好熱菌Thermus thermophilus HB8由来のRbfAの結晶構造を決定し,さらに30S-RbfA複合体の超低温電子顕微鏡像を得ることに成功した.この一連の解析から,16S rRNAが大きな構造変化を起こしていることが判明し,複合体の結晶構造から,世界で初めてRbfAの詳細な結合部位が解明された.決定した構造から,RbfAはmRNAとtRNAの結合部位を占有するだけでなく,16S rRNAの柔軟性を利用してその高次構造を操作することにより,未成熟30Sサブユニットが翻訳を開始しないように制御していることが明らかとなった.
本研究では,高度好熱菌Thermus thermophilus HB8由来のRbfAの結晶構造を決定し,さらに30S-RbfA複合体の超低温電子顕微鏡像を得ることに成功した.この一連の解析から,16S rRNAが大きな構造変化を起こしていることが判明し,複合体の結晶構造から,世界で初めてRbfAの詳細な結合部位が解明された.決定した構造から,RbfAはmRNAとtRNAの結合部位を占有するだけでなく,16S rRNAの柔軟性を利用してその高次構造を操作することにより,未成熟30Sサブユニットが翻訳を開始しないように制御していることが明らかとなった.
PDBj:
2DYJ
PubMed:
17996707
●
翻訳伸長因子EF-G
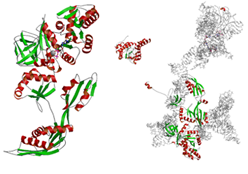
翻訳伸長因子(Elongation Factor: EF)とは,ポリペプチド鎖(タンパク質)が伸長するときに触媒として作用するタンパク質であり,EF-Gは,GTP加水分解酵素である.X線結晶構造解析の結果,GTPが結合したEF-Gのスイッチ領域を含む全長構造の決定に初めて成功した.さらに,X線結晶構造解析から得られた精密な構造情報を電子顕微鏡像にモデリングし,GTPを結合したEF-Gとリボソームの相互作用について詳しい解析を行った.解析の結果,リボソーム30SサブユニットとEF-Gの相互作用を新たに発見し,EF-Gがリボソームの助けを借りて活性型になることを解明した.
PubMed:
17349960
●
tRNAに依存したアミノ酸認識を行うグルタミルtRNA合成酵素
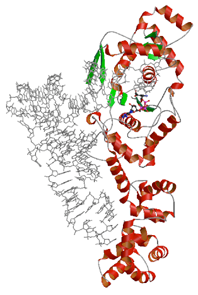
グルタミルtRNA合成酵素(GluRS)とtRNAとの複合体の詳細な比較から,RNAに依存したアミノ酸認識のメカニズムが明らかになった.(1)RNAの結合によってGluRSに構造変化が引き起こされ,結合ポケットの再配置が行われる.(2)RNAの一部が直接グルタミン酸の認識に関わっており,タンパク質と協調して特異的な結合ポケットをかたちづくっていた.RNAとタンパク質が一体となることによってはじめて特異的なアミノ酸認識が達成され,アミノ酸とtRNAの正確な対応づけが可能になるメカニズムが理解された.本研究の成果は,アミノアシルtRNA合成酵素(aaRS)による厳密な基質認識機構を解明し,遺伝暗号翻訳のメカニズムの一端を明らかにしただけでなく,タンパク質とRNAが協調的にはたらくようすを原子レベルの構造から説明した重要なモデルケースである.GluRSのように機能の一部をRNAに依存しているaaRSは,GluRSを含めて3種類しかない.これは生命の起源においてRNA(ないしRNP)がタンパク質合成の主役をつとめていたことの痕跡ととらえることもでき,RNA(やRNP)から多彩な機能をもったタンパク質へと分子の役割が移り変わってきた道筋を考えるうえでも重要な知見となる.
PubMed:
17161369
●
リボソーム30Sサブユニットと抗生物質カスガマイシンとの複合体
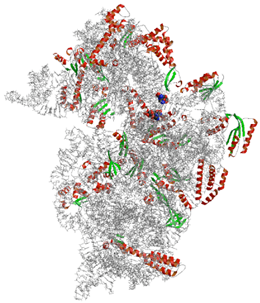
カスガマイシンは,奈良市の春日大社で発見・単離された放線菌が産生する抗生物質で,カビや微生物には効力がありながら,動植物や人体には毒性が低く,農業分野で広く利用されている.リボソーム30Sサブユニットにカスガマイシンが結合した複合体の結晶構造を高分解能で決定したところ,従来予想されていた結合様式とは異なっていることが判明した.2分子のカスガマイシンが30Sサブユニット上のmRNAが結合する部位を占有しており,生化学的な実験の結果とあわせて,カスガマイシンは,tRNAではなくmRNAと30Sサブユニットの相互作用を妨げることにより「翻訳」の開始過程を阻害することを初めて明らかにした.
この作用機序は,これまで明らかにされてきた抗生物質のリボソームでのはたらきとは異なる新規なメカニズムであり,この立体構造解析に基づいた新規薬剤開発が進むと,抗がん剤などの医療用抗生物質創製や農業などの産業応用に結びつくことが期待される.さらに,さまざまな生物のリボソームの構造機能解析を進めることにより,生物種を問わず保存されている部位に結合するカスガマイシンが,動植物に対して毒性が低く,カビや微生物にのみ有効である理由を解明できる可能性が示唆された.
この作用機序は,これまで明らかにされてきた抗生物質のリボソームでのはたらきとは異なる新規なメカニズムであり,この立体構造解析に基づいた新規薬剤開発が進むと,抗がん剤などの医療用抗生物質創製や農業などの産業応用に結びつくことが期待される.さらに,さまざまな生物のリボソームの構造機能解析を進めることにより,生物種を問わず保存されている部位に結合するカスガマイシンが,動植物に対して毒性が低く,カビや微生物にのみ有効である理由を解明できる可能性が示唆された.
PDBj:
2HHH
PubMed:
16998488
●
生命の遺伝暗号表を構築するロイシルtRNA合成酵素(LeuRS)とロイシン用tRNAとの複合体
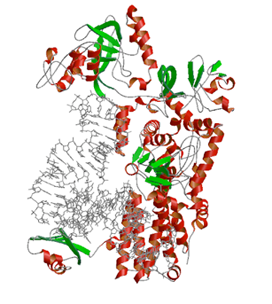
ロイシルtRNA合成酵素(LeuRS)は数千回に一回の割合でイソロイシンという,ロイシンによく似たアミノ酸を誤って認識してしまい,ロイシン用tRNAに結合させてしまう.このときLeuRSの「校正反応ドメイン」が「エラー除去装置」としてはたらき,ロイシン用tRNAに誤って結合したイソロイシンを切り離す.LeuRSがロイシン用tRNAと結合した状態での複合体立体構造を決定した結果,LeuRSは「C末端ドメイン」と呼ばれる部分で,ロイシン用tRNAに特徴的な突き出た長い「可変アーム」の先端を認識していることが分かった.またtRNAの結合の仕方が微妙に異なる2つの複合体構造を明らかにすることができ,これらの2つの状態間では,tRNA中の「認識決定部位」が動いており,異なる様子でLeuRS に結合していた.認識決定部位の動きが,誤って結合したイソロイシンが校正反応ドメインへと運ばれるのかどうかを決めるスイッチとなっており,エラーが起きたときのみこのスイッチがOFFからONに切り替わり,LeuRSはエラーを除去装置へと運ぶモードに切り替わることが明らかになった.
観測された原子構造をもとに,LeuRSとロイシン用tRNAの結合様式を邪魔するような化合物を設計することができれば,それは生物の遺伝暗号表を破綻させることによりはたらく新規の抗生物質として有用となる.またLeuRSのアミノ酸やtRNAの結合部分を人工的にデザインすれば,新しい遺伝暗号表を構築して新規のアミノ酸をタンパク質に導入することも可能である.
観測された原子構造をもとに,LeuRSとロイシン用tRNAの結合様式を邪魔するような化合物を設計することができれば,それは生物の遺伝暗号表を破綻させることによりはたらく新規の抗生物質として有用となる.またLeuRSのアミノ酸やtRNAの結合部分を人工的にデザインすれば,新しい遺伝暗号表を構築して新規のアミノ酸をタンパク質に導入することも可能である.
PubMed:
16155583
●
転写調節因子(DksA)
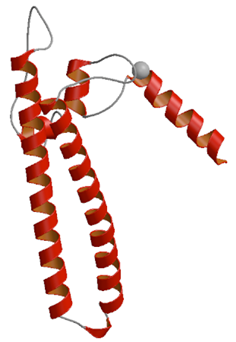
細菌内では,アラーモン(警告物質)と呼ばれるグアノシン4リン酸(ppGpp)がRNAポリメラーゼ(RNAP)の活性中心近傍に結合し,DNA の遺伝情報をRNAへ転写する転写反応を制御する.本研究では,DksAというタンパク質がppGppに依存する転写調節に重要な役割を果たしていることを明らかにした.
大腸菌由来のDksAタンパク質の結晶構造を決定したところ,球状ドメインとコイルドコイルドメインの2つのドメインからなっており,コイルドコイルドメインを形成する2つのα-へリックスの連結部にはアスパラギン酸残基が2つ存在していた.この構造は,RNAポリメラーゼのRNA分解反応を促進する因子「GreA」に類似していた.DksAのコイルドコイルドメインはRNAPの基質侵入孔に入り,ppGppに結合しているマグネシウムイオンとDksAのアスパラギン酸残基とが結合し,ppGpp?RNAP複合体を安定化させていると考えられる.DksAが(GreAとは作用機序が異なるものの)転写伸長反応に影響を及ぼすこと,転写開始反応に対するppGppの効果を増大させること,及び,RNAPに直接結合することによってアスパラギン酸残基がRNAPの活性中心近傍に位置することを生化学的解析によって裏付けた.アスパラギン酸残基を他の残基へ置換した変異DksAには,それらの効果が認められない.
本研究成果は,RNAPの基質侵入孔が複数の転写因子による制御作用のための大事な孔であることを示唆する.転写調節因子に関するタンパク質の解明という学術的な貢献の他,今後の新たな創薬開発に大きく寄与すると考えられる.
大腸菌由来のDksAタンパク質の結晶構造を決定したところ,球状ドメインとコイルドコイルドメインの2つのドメインからなっており,コイルドコイルドメインを形成する2つのα-へリックスの連結部にはアスパラギン酸残基が2つ存在していた.この構造は,RNAポリメラーゼのRNA分解反応を促進する因子「GreA」に類似していた.DksAのコイルドコイルドメインはRNAPの基質侵入孔に入り,ppGppに結合しているマグネシウムイオンとDksAのアスパラギン酸残基とが結合し,ppGpp?RNAP複合体を安定化させていると考えられる.DksAが(GreAとは作用機序が異なるものの)転写伸長反応に影響を及ぼすこと,転写開始反応に対するppGppの効果を増大させること,及び,RNAPに直接結合することによってアスパラギン酸残基がRNAPの活性中心近傍に位置することを生化学的解析によって裏付けた.アスパラギン酸残基を他の残基へ置換した変異DksAには,それらの効果が認められない.
本研究成果は,RNAPの基質侵入孔が複数の転写因子による制御作用のための大事な孔であることを示唆する.転写調節因子に関するタンパク質の解明という学術的な貢献の他,今後の新たな創薬開発に大きく寄与すると考えられる.
PDBj:
1TJL
PubMed:
15294156
●
RNAの分子擬態をしている翻訳伸長因子P (EF-P)
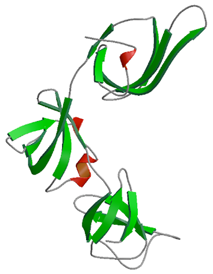
生命の生存に必須であり,遺伝暗号の翻訳過程において重要な因子であるとされながらも,機能や構造の多くの点で謎につつまれたタンパク質である翻訳伸長因子P (EF-P: Elongation Factor P)の立体構造を決定した.EF-Pは3つのドメインからできており,全体の構造はL字型で,トランスファーRNA(tRNA)に酷似していることが明らかになった.本研究で明らかになったEF-Pの立体構造は,翻訳因子はその分子構造をtRNAに擬態させているという“tRNA 擬態”と名付けられた概念を強く支持している.タンパク質であるEF-Pが核酸であるtRNA様の立体構造を模倣することにより,共通の分子基盤をもち,tRNAによって司られている役割(例:遺伝暗号の翻訳など)を果たすのではないかと考えられる.これまでに多くの抗生物質が翻訳過程をターゲットにして作られていることから,本研究の成果が今後の新たな創薬開発に寄与することが期待される.
PDBj:
1UEB
PubMed:
15210970
●
ヒトDNA組換えタンパク質ダブルリング構造(Dmc1)

相同DNA組換え反応において中心的な役割を果たすDmc1の立体構造を決定した. 解析の結果から,Dmc1は,8個のユニットが規則的に円状に並んでリング構造を形成し,さらにそのリング同士が重なって16個のユニットからなる巨大なダブルリング構造をしていることが明らかになった.そして解明された立体構造に基づいた生化学的解析によってDmc1による相同DNA組換え作用機構の解明を試みた.本成果によって,今まで不明であった減数分裂期の相同DNA組換え分子機構の核心部分が明らかになった.Dmc1は,相同DNA組換えに非常に重要なタンパク質であり,本研究成果を利用して細胞内でのDNA組換え効率を人為的に増進させることが可能になれば,今後,染色体上での遺伝子治療の基礎技術など,医療や産業の分野に大きく貢献することが期待される.
PDBj:
1V5W
PubMed:
15125839
●
転移RNAの第二の形(アーケオシンtRNAグアニントランスグリコシラーゼ(ArcTGT)とtRNAとの複合体)

アーケオシンtRNAグアニントランスグリコシラーゼ(ArcTGT)は,古細菌tRNAの修飾塩基であるアーケオシン(7-formamidino-7-deazaguanosine)のtRNAへの導入に関わる修飾酵素である.ArcTGTの標的サイトはtRNAのコア構造に埋もれており,通常のL字型状態のtRNAコアの構造では,ArcTGTは標的サイトにアクセスすることが出来ない.従って,ArcTGTは何らかの構造変化を起こしたtRNAに結合し,修飾塩基を導入していると考えられていたが,具体的なメカニズムは不明であった.
本研究ではArcTGT・tRNA複合体の X線結晶構造解析の結果,ArcTGTは大きく構造変化を起こしたtRNAに結合するとともに,tRNAが修飾の標的部分が露出するように組み変わった「ラムダ(λ)型」構造をとっていることを明らかにした.また,tRNAがL字型に折り畳まっていく段階では,このラムダ型が特に重要であることが明らかになった.本研究で解明された,転移RNAの構造が完成していく仕組みは,このようにさらに複雑なRNAが折り畳んでいく過程への理解への基盤になると考えられる.
本研究ではArcTGT・tRNA複合体の X線結晶構造解析の結果,ArcTGTは大きく構造変化を起こしたtRNAに結合するとともに,tRNAが修飾の標的部分が露出するように組み変わった「ラムダ(λ)型」構造をとっていることを明らかにした.また,tRNAがL字型に折り畳まっていく段階では,このラムダ型が特に重要であることが明らかになった.本研究で解明された,転移RNAの構造が完成していく仕組みは,このようにさらに複雑なRNAが折り畳んでいく過程への理解への基盤になると考えられる.
PDBj:
1J2B
PubMed:
12732145
●
ハエの脚分化を規定する転写因子とDNA の3者複合体の機能解析・構造解析
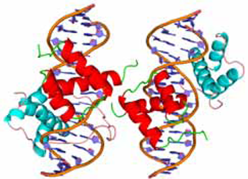
ショウジョウバエの肢の発生過程に働く転写因子間の相互の発現制御機構の解析により,Aristaless (Al) とClawless (Cll)が発現している最先端部でBar が発現しないのは,Al とCll が協調的に働いてBar の発現を抑制しているためであることが明らかになった.Al 単独や,Cll単独では認識 DNA 配列との結合力が極めて低いが,Al-Cll-DNAの三者が共存する条件では強いDNA 結合活性を示す.このような特徴的な結合力の調節を担う構造学的な特徴を明らかにするために,Al と Cll およびそのヒトホモログ CART1(Al ホモログ),Hox11L1(Cll ホモログ) と認識DNA との三者複合体立体構造解析を行い,それぞれの立体構造を決定した.
PubMed:
20389279
●
グルタミンアミドトランスフェラーゼ
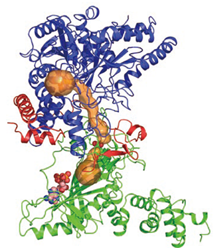
真正細菌や古細菌ではグルタミンをグルタミン用のtRNAに結合させる酵素が存在しない.しかし,グルタミン酸をtRNAに結合させる酵素が,本来の相手ではないグルタミン用のtRNAに結合させる機能を持つ.グルタミン酸がグルタミン用のtRNAに結合したところで,真正細菌ではGatCAB,古細菌ではGatDEという酵素がはたらく.これらの酵素は,グルタミン酸にアンモニアを付け加えてグルタミンに変換する.つまり,グルタミンをそのまま結合させることはできないので,一度グルタミン酸を結合させてから変換し,正しい組み合わせにするわけである.
GatCAB, GatDEそれぞれの構造を解析することにより,グルタミン酸とアンモニアを反応させるしくみが明らかになった.この成果は,新しいアミノ酸を人工的にタンパク質に組み込む技術の開発につながると考えられる.
GatCAB, GatDEそれぞれの構造を解析することにより,グルタミン酸とアンモニアを反応させるしくみが明らかになった.この成果は,新しいアミノ酸を人工的にタンパク質に組み込む技術の開発につながると考えられる.
●
基本転写因子TFIIEα
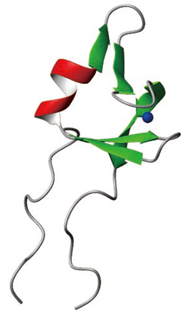
DNAの鎖の上にはタンパク質をつくるための遺伝子がとびとびに並んでおり,細胞がタンパク質をつくるためには,まず遺伝子の部分を見分けて,その部分をもとにmRNAをつくらなければならない.これが転写である.遺伝子の少し手前には,転写を始めるための目印となる部分が存在している.
この目印に結合するのが,基本転写因子というタンパク質である.このタンパク質は何種類もあり,次々にDNAに結合して複合体をつくる.転写を行うのはRNAポリメラーゼという酵素だが,基本転写因子が,転写を開始する位置にRNAポリメラーゼを引き寄せたり,DNAの二重らせんを開いて転写を始められるようにしたりしている.
基本転写因子の一つであるTFIIEαというタンパク質の構造をNMRによって解析し,複合体形成に重要な部分の構造を決定した.この構造の一部のアミノ酸を変えることにより,もとのタンパク質よりも作用の強いタンパク質をつくることにも成功した.これらは,細胞の重要な活動であるタンパク質生成の初期段階のメカニズムの解明に大きく貢献する成果である.
この目印に結合するのが,基本転写因子というタンパク質である.このタンパク質は何種類もあり,次々にDNAに結合して複合体をつくる.転写を行うのはRNAポリメラーゼという酵素だが,基本転写因子が,転写を開始する位置にRNAポリメラーゼを引き寄せたり,DNAの二重らせんを開いて転写を始められるようにしたりしている.
基本転写因子の一つであるTFIIEαというタンパク質の構造をNMRによって解析し,複合体形成に重要な部分の構造を決定した.この構造の一部のアミノ酸を変えることにより,もとのタンパク質よりも作用の強いタンパク質をつくることにも成功した.これらは,細胞の重要な活動であるタンパク質生成の初期段階のメカニズムの解明に大きく貢献する成果である.
PDBj:
1VD4
PubMed:
15385556
●
細胞内輸送タンパク質(GGA)
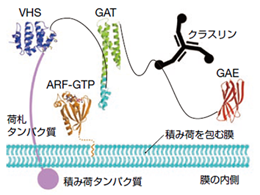
細胞の中では,つねにダイナミックな活動が行われている.細胞の重要な活動の一つとして,ものの輸送がある.細胞内には,膜で包まれたさまざまな小器官があり,それらの間でもののやりとりが活発に行われる.
その際,運ばれるものも,膜に包まれた状態で細胞内を移動する.まちがいなく運ぶために,膜の外には積み荷に応じた荷札のタンパク質が突き出しており,別のタンパク質がこの荷札を見分け,積み荷を目的の場所へと運んでいく.
このような運搬人の機能をもつタンパク質の一つとして,GGAが知られている.細胞の中には,ゴルジ体という,平べったい袋が積み重なったような小器官があり,その端がくびれきれるようにして,積み荷の入った袋ができる.GGAがこの積み荷をリソソーム(不要になったものを分解する小器官)まで運ぶ.
GGAは3つの部分からなり,それぞれの部分が別々のタンパク質と作用することで,積み荷が目的の場所まで運ばれるようにしている.3つの部分のそれぞれについて構造を決定することにより,まちがいのない物質輸送のメカニズムが明らかとなってきた.この成果は,細胞の活動を探る上でも,細胞内での輸送異常が原因の病気の治療薬開発においても,貢献すると期待される.
その際,運ばれるものも,膜に包まれた状態で細胞内を移動する.まちがいなく運ぶために,膜の外には積み荷に応じた荷札のタンパク質が突き出しており,別のタンパク質がこの荷札を見分け,積み荷を目的の場所へと運んでいく.
このような運搬人の機能をもつタンパク質の一つとして,GGAが知られている.細胞の中には,ゴルジ体という,平べったい袋が積み重なったような小器官があり,その端がくびれきれるようにして,積み荷の入った袋ができる.GGAがこの積み荷をリソソーム(不要になったものを分解する小器官)まで運ぶ.
GGAは3つの部分からなり,それぞれの部分が別々のタンパク質と作用することで,積み荷が目的の場所まで運ばれるようにしている.3つの部分のそれぞれについて構造を決定することにより,まちがいのない物質輸送のメカニズムが明らかとなってきた.この成果は,細胞の活動を探る上でも,細胞内での輸送異常が原因の病気の治療薬開発においても,貢献すると期待される.
2,
創薬基盤研究
●
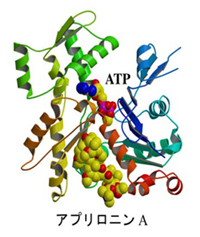
アクチン/アプリロニンA 複合体
●
マウスカルノシナーゼ
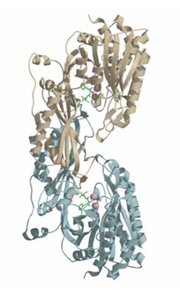
カルノシナーゼ(カルノシン加水分解酵素)はジペプチドであるL-カルノシン(β-alanyl-L-histidine)を加水分解するペプチダーゼであり,ほ乳類などの脳などに分布している.L-カルノシンは,生理活性のあるジペプチドで,ほ乳類などの中枢神経系に存在し,エネルギー恒常性維持の制御や神経保護に関わっている.
カルノシナーゼには性質が違うCN1とCN2があり,今回,マウスCN2とインヒビターであるbestatinとの複合体のX線結晶解析を行った.CN2は2個金属酵素であり,Zn酵素,Mn酵素を調製し,それぞれ独立にMAD法(多波長異常分散法)で結晶解析を行った.
カルノシナーゼには性質が違うCN1とCN2があり,今回,マウスCN2とインヒビターであるbestatinとの複合体のX線結晶解析を行った.CN2は2個金属酵素であり,Zn酵素,Mn酵素を調製し,それぞれ独立にMAD法(多波長異常分散法)で結晶解析を行った.
PubMed:
18550540
●
Trypanosoma brucei 由来PGF合成酵素のX線解析とNADPHによる還元反応の機構解明
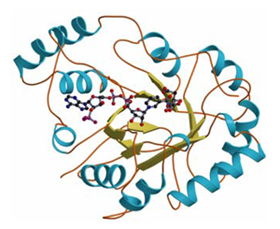
プロスタグランジン(PG)F2αは,PGF合成酵素の働きによって生合成され,子宮平滑筋の収縮作用を有し,陣痛促進剤として広く臨床応用されている生理活性物質である.しかし,寄生虫に感染した宿主の血中に検出可能なレベルでPGF2αが生合成され,Nagana病に見られるような家畜の流産などが深刻化している.そこでNagana病の原因となる原虫トリパノソーマTrypanosoma bruceiが持つヒトと薬剤感受性の異なるPGF合成酵素を分子生物学的に同定し,2.1Å分解能で構造を決定した.本酵素はアルドケト還元酵素(AKR)のスーパーファミリーに属し,活性中心にはCatalytic tetradと呼ばれる4つのアミノ酸残基が存在し,どのアミノ酸が欠損しても活性がなくなることが示唆された.しかし,Mutation実験を行い,Lys77およびHis110の2個のみが反応に関与し,Catalytic dyadで進行する新たな反応機構を提唱した.
PDBj:
1VBJ
PubMed:
15845552
●
多剤排出ポンプ調節因子の構造解析
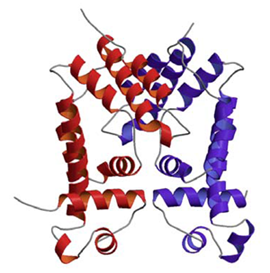
CGL2612タンパク質はC.glutamicumのゲノム解析により新たに見いだされた転写因子である.立体構造解析の結果,本タンパク質はアミノ酸配列の相同性の低さにも関わらずS.aureus (ブドウ球菌)由来の多剤耐性因子QacRと非常に似た立体構造を持つことが明らかとなった(一次構造の相同性は7%).またSELEX法を用いたCGL2612による結合DNA配列決定実験により,major facilitator superfamilyに属するポンプタンパク質をコードしていると予想されるcgl2611遺伝子の発現調節を行うことがわかった.実際,cgl2612破壊株では種々の薬剤に対して感受性が低下することが明らかとなった.これらのことよりCGL2612は転写抑制因子として働き,薬剤排出ポンプCGL2611と組となってC.glutamicumの薬剤耐性化に関わっていることが示唆された.C.glutamicumはジフテリア菌や結核菌の類縁菌であるので,CGL2612タンパク質の構造は,これら病原菌の多剤耐性因子のモデルとなりうる.
PubMed:
16166084
●
SARS プロテアーゼ(3CL-PRO)
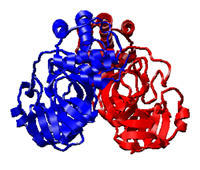
大腸菌無細胞系を用い,自己プロセシングによる活性化プロセスを再現し,天然の活性型と同一のアミノ酸配列を持つターゲットタンパク質の大量発現と精製に初めて成功した.そのX線結晶構造解析により,発現された全領域での高分解能(1.7Å)の立体構造決定に成功した.すでにホモロジーモデルに対するin silicoスクリーニングで得ている阻害化合物を,本高分解能活性型構造を用いてさらに最適化している.
PDBj:
2DUC
●
Keap 1
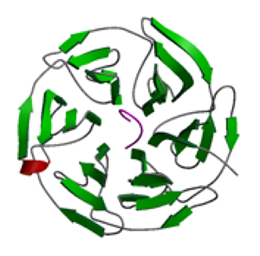
X線結晶構造解析により,Keap1のダブルグリシンリピート領域とC末端領域(Keap1-DC)の立体構造を決定することにより,Keap1-DCがNrf2と結合するために特徴的なリング複合体を形成していることを明らかにした.さらに,肺がん細胞においてKeap1-DCに生じる2ヶ所のアミノ酸変異がNrf2に対する結合能を低下させることを構造レベルで明らかにした.酸化ストレスの生体内センサーとして働くKeap1と遺伝子発現活性化に働く転写因子Nrf2とが形成する複合体の構造を初めて決定したことで,酸化ストレスに対する生体防御機構の分子レベルの理解を進めた.また,肺がん症例において,Keap1の機能に変換をもたらす複数の遺伝子変異が存在することを見出し,その機能変換メカニズムの構造的基盤を明らかにした.酸化ストレス応答機構のメカニズムの破綻による疾患発症との相関性の存在についての理解に重要な知見を与えたことは,癌研究をはじめとした臨床医学的にも非常に興味深く,今後の更なる研究展開が期待される.
●
ヒト,プロスタグランジンD 合成酵素

炎症・アレルギー反応のmediatorとなるProstaglandin D2 (PGD2)をPGH2から合成するヒト由来hematopoietic protstaglandin D synthetase (H-PGDS)は,抗アレルギー・抗炎症薬の重要ターゲットとなっている.炎症・アレルギー反応のmediatorとなるProstaglandin D2 (PGD2)受容体にはCRTH2とDPの2種類が知られているため,PGD2の阻害剤としては受容体アンタゴニストよりもPGD合成酵素を阻害するブロッカーの開発の方が容易であると考えられている.
このH-PGDSを結晶化してX線結晶構造解析を実施し,2量体の中央にCaあるいはMgイオンの結合部位を持つ構造の座標情報を明らかにした.更に,脳損傷や筋ジストロフィーの症状を有するマウスへの経口投与で効果のある阻害剤との共結晶化および複合体のX線結晶構造解析に成功し,この構造情報を基に,更なるStructure-based Drug Designを展開してより活性の高い誘導体を開発しており,一部,複合体構造を明らかにした.
このH-PGDSを結晶化してX線結晶構造解析を実施し,2量体の中央にCaあるいはMgイオンの結合部位を持つ構造の座標情報を明らかにした.更に,脳損傷や筋ジストロフィーの症状を有するマウスへの経口投与で効果のある阻害剤との共結晶化および複合体のX線結晶構造解析に成功し,この構造情報を基に,更なるStructure-based Drug Designを展開してより活性の高い誘導体を開発しており,一部,複合体構造を明らかにした.
●
ペプチジルアルギニンデイミナーゼ4
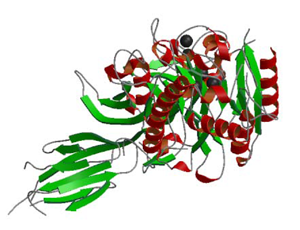
Peptidyl arginine deiminase (PAD)は,Caイオン依存的にアルギニンをシトルニンに変換する酵素である.PAD4はヒストン中のアルギニンのメチル化を制御しているタンパク質で,好酸球や好中球で発現している.
今回の研究で,PAD4の立体構造やこれまでにない全く新しいCaイオンによる活性化機構を解明した.更に関節リウマチ(RA)発病者の血液中にはシトルリン化されたタンパク質に対する自己抗体ができることから,PAD4がRAの発症と強い相関があると考えられ,PAD4阻害剤がRAの治療薬になる可能性が有ると期待されている.
今回の研究で,PAD4の立体構造やこれまでにない全く新しいCaイオンによる活性化機構を解明した.更に関節リウマチ(RA)発病者の血液中にはシトルリン化されたタンパク質に対する自己抗体ができることから,PAD4がRAの発症と強い相関があると考えられ,PAD4阻害剤がRAの治療薬になる可能性が有ると期待されている.
PubMed:
15247907
●
リポタンパク質局在化因子(LolA, LolB)
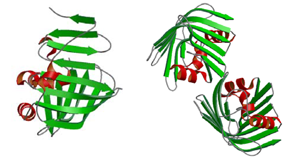
細菌は自らの形態を維持し,生体内の諸物質を輸送し,また薬剤を排出する為に,何十種類ものリポタンパク質を持っている.グラム陰性菌にある二重の生体膜(内膜と外膜)間のペリプラズム空間でのリポタンパク質の輸送をつかさどる一連のタンパク質群として,ペリプラズム空間のシャペロンタンパク質LolA,外膜でのリポタンパク質受容体LolBなどが知られている.これらのタンパク質の欠損は細菌にとって致命的であり,グラム陰性菌の生存に本質的な役割を果たしている.
LolA,LolBの立体構造を初めて明らかにしたことにより,タンパク質輸送の更なる解明や,立体構造を基にした病原性のグラム陰性菌(O-157大腸菌,サルモネラ菌等)を特異的ターゲットとする抗生物質の開発につながることが期待される.
LolA,LolBの立体構造を初めて明らかにしたことにより,タンパク質輸送の更なる解明や,立体構造を基にした病原性のグラム陰性菌(O-157大腸菌,サルモネラ菌等)を特異的ターゲットとする抗生物質の開発につながることが期待される.
PubMed:
12839983
●
Brd2 ブロモドメインBD1/ヒストンH4 テイル複合体
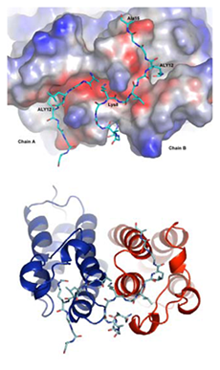
遺伝子発現が活性化されたヒト細胞は,染色体構成タンパクであるヒストンH4の12番目のリジン残基が恒常的にアセチル化される特徴を持っている.Brd2のブロモドメインは,このアセチル化されたリジン残基と結合して,近隣に位置する遺伝子の発現を活性化することが知られていたが,その制御メカニズムは不明であった.本研究では,Brd2のブロモドメイン(BD1)と,ヒストンH4のアセチル化されたテイル領域との複合体の立体構造を初めて明らかにした.その結果,Brd2-BD1は二量体を形成することにより,本来特定の構造を作りにくいヒストンH4テイルに15残基分の構造を形成させ,その12 番目のアセチル化リジン残基を特異的に認識する機構を明らかにした.この研究は,染色体の活性化/不活性化に関わるエピジェネティックな情報(単なる遺伝情報を越えた上位階層の情報:ヒストン・コード)を維持/発現する機構を構造的に実証した点で意義がある.
またBRD2遺伝子は,全てんかん患者の約8%を占める若年性ミオクロニーてんかん(JME)患者家系で変異が生じていることが報告されており,今後,JMEに対する新たな治療法への応用も期待される.
またBRD2遺伝子は,全てんかん患者の約8%を占める若年性ミオクロニーてんかん(JME)患者家系で変異が生じていることが報告されており,今後,JMEに対する新たな治療法への応用も期待される.
PubMed:
20048151
●
キサンチン酸化還元酵素
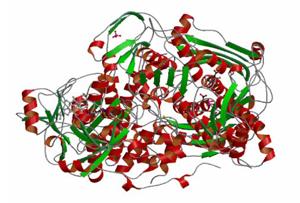
キサンチン酸化還元酵素は,尿酸を産生するプリン代謝経路の最終2段階(ヒポキサンチン→キサンチンおよびキサンチン→尿酸)において機能する.血液中の尿酸の濃度が上昇すると,結晶化して足の親指付け根などの関節にたまり,痛風発作と呼ばれる炎症を起こす.本酵素の阻害剤は,痛風治療薬として知られている.
本研究では,薬物との複合体構造を解析して,創薬活動に重要なデータを得ると共に,本酵素の活性酸素産生能に注目して研究を進めている.活性酸素を介したシグナル伝達機構は,発生・分化に重要な意義を持つと考えられる.また,活性酸素を大量に生産する変異体酵素を作成し,本変異体遺伝子をノックインしたマウスは疾患及び老化モデル生物として医学,薬学分野での幅広い利用が期待できる.
本研究では,薬物との複合体構造を解析して,創薬活動に重要なデータを得ると共に,本酵素の活性酸素産生能に注目して研究を進めている.活性酸素を介したシグナル伝達機構は,発生・分化に重要な意義を持つと考えられる.また,活性酸素を大量に生産する変異体酵素を作成し,本変異体遺伝子をノックインしたマウスは疾患及び老化モデル生物として医学,薬学分野での幅広い利用が期待できる.
●
「メラニン色素」の輸送に必須なタンパク質Rab27複合体
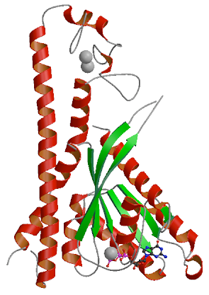
Rab27Aは,Slac2-a(別名メラノフィリン:Melanophilin),Slp2-a(別名エキソフィリン4:Exophilin4)という2つのエフェクターを連続的に用いて,メラノソームをメラノサイトの核周辺から細胞膜まで輸送する.本研究では,メラニン色素輸送に中心的な役割を果たしている2つのタンパク質複合体「Rab27A‐Slp2-a」および「Rab27B‐Slac2-a」の構造を明らかにした.
解明した構造情報から,Rab27とSlac2-aおよびSlp2-aとの結合に重要なアミノ酸残基を同定し,Rab27が特定のエフェクターを認識する基盤を解明するとともに,Rab27AとSlac2-aのアミノ酸残基の変異が,肌の白色化の症状を示すグリセリ(Griscelli)症候群という遺伝病を発症する分子メカニズムを明らかにした.本研究成果は,これらのタンパク質間の結合を阻害あるいは安定化するような薬剤の設計に応用することが可能であり,メラノソーム輸送を人為的に制御することによって,肌の美白の維持や白髪発生の抑制につながる可能性が期待される.
解明した構造情報から,Rab27とSlac2-aおよびSlp2-aとの結合に重要なアミノ酸残基を同定し,Rab27が特定のエフェクターを認識する基盤を解明するとともに,Rab27AとSlac2-aのアミノ酸残基の変異が,肌の白色化の症状を示すグリセリ(Griscelli)症候群という遺伝病を発症する分子メカニズムを明らかにした.本研究成果は,これらのタンパク質間の結合を阻害あるいは安定化するような薬剤の設計に応用することが可能であり,メラノソーム輸送を人為的に制御することによって,肌の美白の維持や白髪発生の抑制につながる可能性が期待される.
PDBj:
2ZET
PubMed:
18940604
●
肝臓がん原因タンパク質ガンキリンとタンパク質分解を行うサブユニットとの複合体構造
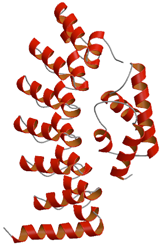
ガンキリンは肝臓がんの原因遺伝子産物として発見され,細胞周期の制御機構に関与する様々なタンパク質に作用し,肝臓がん細胞の異常増殖を引き起こすことが知られている.本研究では,このがんタンパク質であるガンキリンと,ユビキチン化されたタンパク質を分解する26SプロテアソームのサブユニットであるS6ATPaseドメインとの複合体について立体構造を解明し,ガンキリンとS6ATPaseとの結合部位を明らかにした.さらに,ガンキリンが分解を促進するタンパク質で,がん抑制タンパク質の1つでもあるpRbとガンキリンの結合部位は既に明らかとなっているが,S6ATPaseとの結合部位の一部は,そのpRbの結合部位と同じであったことから,ガンキリンがpRbの分解を促進するメカニズムについての新規モデルを提案した.本研究に基づいた阻害剤開発は,ガンキリンに依存したがん細胞の異常増殖を抑える,新規抗がん治療薬の開発につながると期待される.
PDBj:
2DVW
PubMed:
17292836
●
腫瘍壊死因子のシグナルが細胞の核内に伝達される際に重要な役割を果たすCYLDタンパク質
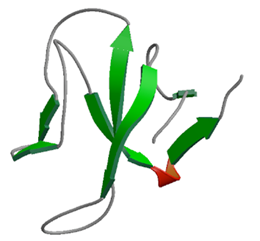
生体内での免疫応答や炎症反応の過程において,腫瘍壊死因子のシグナルが細胞の核内に伝達される際に重要な役割を果たすCYLDタンパク質のシグナル伝達ネットワーク上への関与のメカニズムを明らかにするために,まずIKKと呼ばれるリン酸化酵素複合体の調節サブユニットであるNEMOと相互作用する CYLD内の領域の特定を試みた結果,3番目のCAP-Glyドメインがこの相互作用には必要十分であることが明らかになった.このドメイン領域を無細胞タンパク質合成系により安定同位体で標識して合成し,精製を行い,NMRを用いて立体構造を決定した.タンパク質表面の解析を行ったところ,伸びたペプチド鎖が結合しうる疎水性の溝があることが明らかになった.一方で,相互作用因子であるNEMOのアミノ酸配列を分析し,相互作用部位の候補となりうるプロリンに富む領域を2ヶ所特定した.これらのアミノ酸配列に相当するペプチドを用いた解析の結果,これらのうちの1つと特異的に結合することが明らかになり,NEMO上の結合部分の特定に成功した.さらに NMRを用いた解析により,上記の溝がこの結合面であることが実験的に明らかとなった.本成果によって,TRAFからIKKへのシグナル伝達の分子メカニズムの一端が明らかになった.本成果は腫瘍抑制のメカニズムの理解につながるものであり, 学術的な貢献の他にも今後の新たな創薬ターゲットとしても期待される.
PDBj:
1IXD
PubMed:
15341735
●
モノアミン酸化酵素
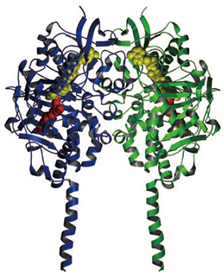
私たちの脳の中には,膨大な数の神経細胞があり,セロトニンやノルアドレナリンなどの神経伝達物質が神経細胞から神経細胞へと情報を伝えている.これらの神経伝達物質の量は,つねに厳密にコンロトールされているが,神経伝達物質が不足して微妙なバランスが崩れると,思考が不活発になったり意欲が低下したりし,うつ病などさまざまな神経疾患を引き起こす.
脳の中で,神経伝達物質を分解して減らすはたらきをしているのは,モノアミン酸化酵素というタンパク質である.このタンパク質のはたらきを阻害すれば,神経伝達物質の減少が抑えられ,多くの神経疾患を治すことができると考えられる.
そこで,モノアミン酸化酵素と阻害物質の複合体の立体構造を解析し,阻害物質が酵素のどこの部分に入り込んでいるかを明らかにした.この知見をもとに,うつ病の新しい治療薬の開発が期待されている.
脳の中で,神経伝達物質を分解して減らすはたらきをしているのは,モノアミン酸化酵素というタンパク質である.このタンパク質のはたらきを阻害すれば,神経伝達物質の減少が抑えられ,多くの神経疾患を治すことができると考えられる.
そこで,モノアミン酸化酵素と阻害物質の複合体の立体構造を解析し,阻害物質が酵素のどこの部分に入り込んでいるかを明らかにした.この知見をもとに,うつ病の新しい治療薬の開発が期待されている.
PDBj:
1O5W
PubMed:
15050826
●
細胞のがん化を促進するタンパク質(CRK)
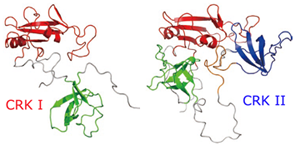
私たちの体は,数十兆個の細胞から構成されている.その数はほぼ一定に保たれ,細胞が増えすぎないように制御するメカニズムがはたらいている.ところが,このメカニズムが正常にはたらかなくなると,細胞は無制限に増え続けてしまい,増えすぎた細胞が他の組織や臓器に転移して機能不全を起こしたり,細胞の増殖に栄養が奪われて身体が衰弱したりするなど,生命の維持に重大な支障をきたす場合がある.これが「がん」である.
がんは,遺伝子の突然変異によりタンパク質が異常なはたらきをしたり,通常の機能を果たせなくなったりすることで引き起こされる.細胞のがん化を促進させるタンパク質の1つにCRKがある.CRKには2つのタイプがあり,CRK I だけが細胞をがん化させる能力をもっている.両者の構造を明らかにし比較することで,細胞のがん化を引き起こすタンパク質の構造的な要因が明らかになった.今後,こうしたタンパク質をターゲットとした抗がん剤などの開発が期待される.
がんは,遺伝子の突然変異によりタンパク質が異常なはたらきをしたり,通常の機能を果たせなくなったりすることで引き起こされる.細胞のがん化を促進させるタンパク質の1つにCRKがある.CRKには2つのタイプがあり,CRK I だけが細胞をがん化させる能力をもっている.両者の構造を明らかにし比較することで,細胞のがん化を引き起こすタンパク質の構造的な要因が明らかになった.今後,こうしたタンパク質をターゲットとした抗がん剤などの開発が期待される.
PubMed:
17515907
3,
酵素化学研究基盤
●
ディールス・アルダー反応触媒酵素の構造解析
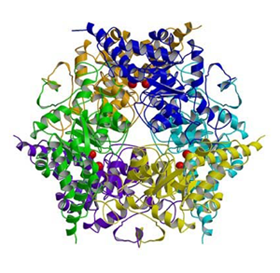
ディールス・アルダー反応は,有機合成化学上極めて重要な反応の一つであり,制癌剤や抗生物質,免疫抑制剤など,芳香族化合物の合成現場で盛んに使用されている.この反応を触媒する酵素の存在は,数十年前より予測されていたが,触媒反応を説明することは容易ではなかったため,酵素の存在を証明した例は無かった.我々は,マクロフォミン酸合成酵素(MPS)と,基質の一つであるピルビン酸との複合体の状態で立体構造を解析することに成功した.得られた立体構造情報および実験データから,世界で初めて天然型Diels-Alderaseの触媒機構を説明することができた.天然型Diels-Alderaseはこれまで創製されていたDiels-Alderase触媒抗体に比べ,精緻な工夫を随所に施し,格段に高速な反応効率性を実現していることが明らかになった.
PDBj:
1IZC
PubMed:
12634789
●
ホスホセリンスルフヒドリル化酵素
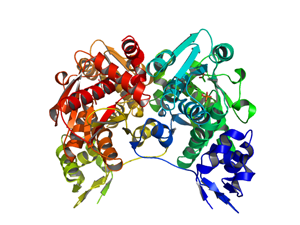
O-phosphoserine sulfhydrylaseは,好気性超好熱性古細菌Aeropyrum pernix K1に見られる新規な酵素である.この酵素は,O-phospho-L-serineとsulfide(-SH-)から,新規なシステイン合成反応を触媒する.
研究面ではこの酵素が従来知られていなかった基質からシステインを合成できること,すなわち知財権に守られたシステイン合成の新規経路を提供できることを示し発表した.新規酵素であることは国際生化学連合が酵素番号EC2.5.1.65を付与することによっても示すことができた.
タンパク3000の成果として,この酵素(システイン合成酵素)の「ホスホセリンに対する基質特異性を高める方法」を別途特許出願した.この出願は,タンパク3000で解明した立体構造の知識を活用し,立体構造がなければ分からなかった特定のアミノ酸残基を改変して基質特異性を変化させる方法である.
研究面ではこの酵素が従来知られていなかった基質からシステインを合成できること,すなわち知財権に守られたシステイン合成の新規経路を提供できることを示し発表した.新規酵素であることは国際生化学連合が酵素番号EC2.5.1.65を付与することによっても示すことができた.
タンパク3000の成果として,この酵素(システイン合成酵素)の「ホスホセリンに対する基質特異性を高める方法」を別途特許出願した.この出願は,タンパク3000で解明した立体構造の知識を活用し,立体構造がなければ分からなかった特定のアミノ酸残基を改変して基質特異性を変化させる方法である.
PDBj:
1WKV
PubMed:
16005886
●
3-ヒドロキシ酪酸脱水素酵素
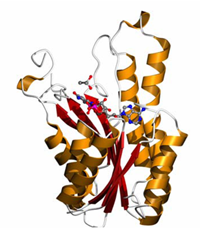
●
γ-グルタミルシステイン合成酵素
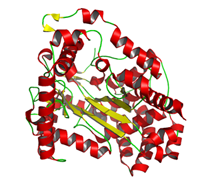
●
制限酵素PabI
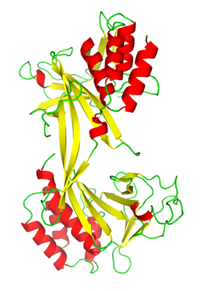
制限酵素PabIは超好熱古細菌Pyrococcus abyssiから発見された新規制限酵素である.PabIは,既存の制限酵素とは全く異なる突出末端を残してdsDNAを切断することができるため,その応用が期待されている.またアミノ酸配列から予測される立体構造も既存の制限酵素とは大きく異なっていることが予測されている.
本研究では,小麦胚芽を用いたタンパク質無細胞発現系を用いPabIを大量調製し結晶化実験を行うことにより,X線結晶構造解析法によってその立体構造を決定することに成功した.その結果,PabIは予測通り新規のフォールドを持つ制限酵素であることが明らかにされた.構造情報を元にPabIのDNA認識部位及び活性に関わるアミノ酸残基の推定が可能となった.PabIは新規の切断部位を持つ重要な制限酵素であるが,立体構造から詳細な反応機構を解明しその酵素活性を最適化すれば,遺伝子工学の分野での更なる応用が期待される.
本研究では,小麦胚芽を用いたタンパク質無細胞発現系を用いPabIを大量調製し結晶化実験を行うことにより,X線結晶構造解析法によってその立体構造を決定することに成功した.その結果,PabIは予測通り新規のフォールドを持つ制限酵素であることが明らかにされた.構造情報を元にPabIのDNA認識部位及び活性に関わるアミノ酸残基の推定が可能となった.PabIは新規の切断部位を持つ重要な制限酵素であるが,立体構造から詳細な反応機構を解明しその酵素活性を最適化すれば,遺伝子工学の分野での更なる応用が期待される.
PDBj:
2DVY
PubMed:
17332011
●
ヨードチロシンを特異的に認識する古細菌由来のチロシルtRNA合成酵素とヨードチロシンの複合体
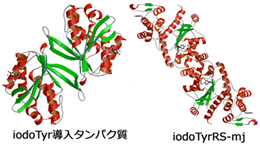
本研究では,大腸菌のシステムによるタンパク質へのヨードチロシン導入を達成すべく,古細菌由来のチロシルtRNA合成酵素(TyrRS)を改変した.様々なTyrRSの変異体作成を行い,ヨードチロシンとだけ結合するiodoTyrRS-mjを作成することに成功した.さらに,この変異体の立体構造を解明し,iodoTyrRS-mjがどのようにしてヨードチロシンを識別しているのかを明らかにした.ヨードチロシンのヨード原子は,チロシン結合ポケットにある5つのアミノ酸残基の間にはまり込むようにして酵素に結合していたが,このような相互作用はヨード原子を持っていないチロシンの場合には見られないため,チロシンがiodoTyrRS-mjとは結合しないことが判明した.さらに,古細菌チロシンtRNAを発現させた大腸菌を用いた,ヨードチロシンの部位特異的なタンパク質導入にも成功した.
PubMed:
19278648
●
校正機能をもったヨードチロシンを特異的に認識するチロシルtRNA合成酵素とヨードチロシン複合体
●
重い原子を含むアミノ酸を認識してtRNAに結合させる変異チロシルtRNA合成酵素
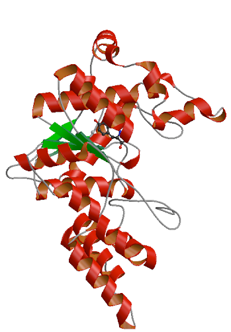
アミノアシルtRNA合成酵素を遺伝子工学的に改変すると,暗号解読表を人為的に書き換えることができる.例えば,天然で使われる20種類のアミノ酸以外の,全く新しいアミノ酸と新しいtRNAを結びつける合成酵素を作れば,新しいアミノ酸を特定の塩基配列に結びつけることができる.2002年に,遺伝子工学によって作り出した変異TyrRSを用いて,ヨウ素という重い原子を含む非天然のアミノ酸を,タンパク質に自由に導入することに成功した.このような非常に重い原子は電磁波に対する性質が違うので,タンパク質の立体構造の決定にこのヨウ素を含む「超」タンパク質を用いることが可能である.実際に変異TyrRSは,鍵穴のようなポケット状のくぼみをもち,ヨードチロシンがちょうど鍵のようにその中にぴったりはまることで,tRNAとヨードチロシンの結合ができることが明らかとなった.
このように,人工的に作られたタンパク質が巧みにヨードチロシンのような非天然の基質を認識する様子を捉えたのは世界でもほとんど類を見ない.観測された原子構造をもとにして,変異TyrRSのアミノ酸結合ポケットをさらに人工的にデザインすることが可能になり,より新しいアミノ酸をうまく認識させることができると期待され,こうして作られた人工的なTyrRSは,工業や医薬,癌などの診断に役立つ超タンパク質の生産という応用性を飛躍的に高めることができると考えられる.
このように,人工的に作られたタンパク質が巧みにヨードチロシンのような非天然の基質を認識する様子を捉えたのは世界でもほとんど類を見ない.観測された原子構造をもとにして,変異TyrRSのアミノ酸結合ポケットをさらに人工的にデザインすることが可能になり,より新しいアミノ酸をうまく認識させることができると期待され,こうして作られた人工的なTyrRSは,工業や医薬,癌などの診断に役立つ超タンパク質の生産という応用性を飛躍的に高めることができると考えられる.
PubMed:
15671170
●
新種タンパク質をつくる鍵となるチロシルtRNA合成酵素
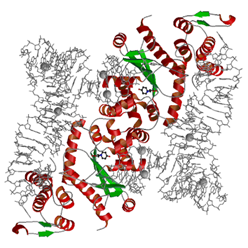
人工的なアミノ酸を組み込んだ新種タンパク質を合成する鍵となる酵素(チロシルtRNA合成酵素 (TyrRS) )の原子分解能での立体構造を決定した.その結果,TyrRSがアミノ酸を深いポケットで識別し.特定のtRNAだけを結合するために, TyrRSがtRNAの塩基を原子レベルで緻密に認識している様子が明らかになった.実際に,観測された原子構造をもとにして,TyrRSのアミノ酸結合ポケットを人工的にデザインすることで,新しいアミノ酸やtRNAを強く認識させることに成功した.本研究で解明された,アミノ酸とtRNA両方の特異性に関わる部分の構造情報を用いれば,あらゆるアミノ酸を酵素の人工的なデザインによって自由に細菌に取り入れさせることが可能になる.例えば,構造情報に基づいた酵素のコンピュータデザインなどによって,工業的に有用な酵素や,がん特効薬などの医薬品となるタンパク質を自由に開発できると期待される.
PDBj:
1J1U,
PubMed:
12754495
●
プロリルペプチダーゼ
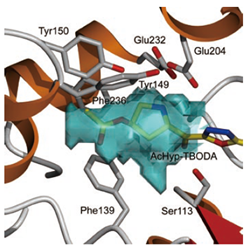
コラーゲンはヒトの体のタンパク質のうち30%近くを占めており,皮膚や内臓,骨などあらゆる組織に存在し,細胞同士をつなぎとめる役割をしている.コラーゲンは3本の鎖が縄のように束になった構造をしているため張力があり,そのおかげで,皮膚は強さや張りが維持され,骨は折れにくくなる.
最近,皮膚にあるコラーゲンは,細菌が体内に侵入するのを防ぐ「よろい」のようなはたらきをしていることもわかってきた.ところが,細菌の中には体内に侵入し,感染を引き起こすものが存在する.こうした細菌はコラーゲンの成分を分解するプロリルペプチダーゼという酵素をもっているため,コラーゲンのバリアを破ることが可能である.
日和見感染症を引き起こす細菌由来のプロリルペプチダーゼと,その機能を阻害する物質の複合体の立体構造の解析により,この酵素の機能を効果的に阻害する物質のしくみがわかってきた.この成果は,細菌感染を防ぐ治療薬の開発につながると期待される.
最近,皮膚にあるコラーゲンは,細菌が体内に侵入するのを防ぐ「よろい」のようなはたらきをしていることもわかってきた.ところが,細菌の中には体内に侵入し,感染を引き起こすものが存在する.こうした細菌はコラーゲンの成分を分解するプロリルペプチダーゼという酵素をもっているため,コラーゲンのバリアを破ることが可能である.
日和見感染症を引き起こす細菌由来のプロリルペプチダーゼと,その機能を阻害する物質の複合体の立体構造の解析により,この酵素の機能を効果的に阻害する物質のしくみがわかってきた.この成果は,細菌感染を防ぐ治療薬の開発につながると期待される.
PubMed:
16452443
